
Лаборатория функциональной геномики и протеомики растений
C 25 по 30 сентября 2023 года во Владивостоке прошла Школа молодых ученых "Новые биотехнологии в функциональной геномике, растениеводстве и защите растений" при поддержке Российского Научного Фонда (проект № 23-74-30003).


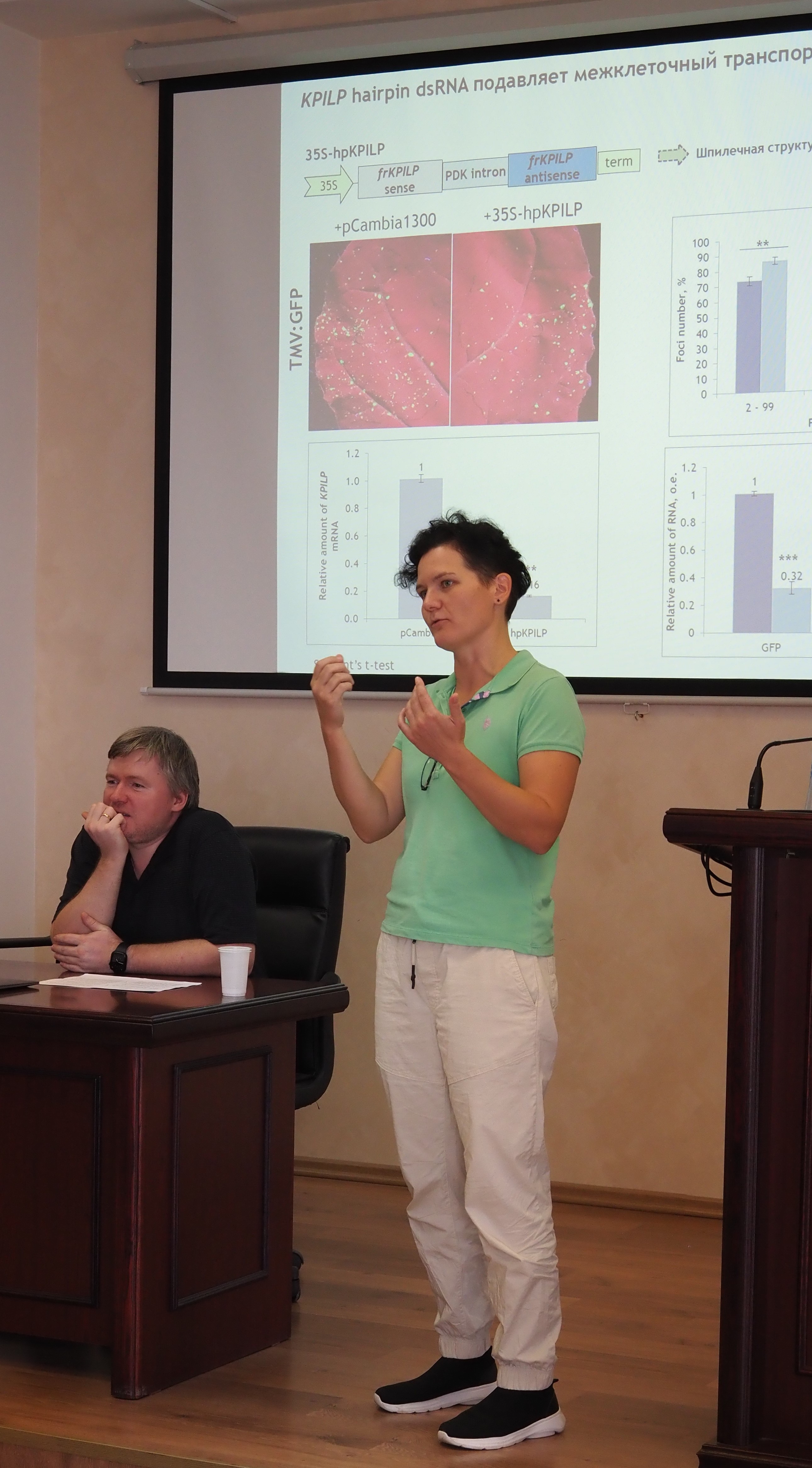

Благодарим всех за сотрудничество и участие в Школе!
Ознакомиться с тезисами участников Школы можно по ссылке: Тезисы
Электронная почта для связи: bio.plants@mail.ru
С уважением, Организационный комитет.
| ФИО | Должность | Контакты |
|---|---|---|
| Тальянский Михаил Эммануилович, проф., д.б.н. | г.н.с. | |
| Борщевская Л.Н. | с.н.с. | |
| Калинина Н.О., д.б.н. | с.н.с. | |
| Комарова Т.В. | с.н.с. | |
| Кост О.А. | с.н.с. | |
| Спеченкова Надежда Андреевна, к.х.н. | н.с. | |
| Попова Е.В. | м.н.с. | |
| Самарская Виктория Олеговна | м.н.с. | |
| Тихомирова В.Е. | м.н.с. | |
| Ильина Ирина Юрьевна | н.с. | |
| Петрова Елена Эдуардовна, к.б.н. | инженер |
 Загрузка...
Загрузка...Научные проекты
Загрузка...Тальянский Михаил Эммануилович
Москва, ул. Миклухо-Маклая, 16/10 — На карте
Загрузка...