
Пресс-центр / новости / Наука /
Восьмая формула света
Феномен биолюминесценции известен около двух с половиной тысячелетий, однако только в ХХ веке ученые вплотную взялись за изучение его химической природы. По современным оценкам, существует около 30 различных биолюминесцентных систем, но на данный момент известны структуры только семи природных люциферинов, последняя из которых была расшифрована 25 лет назад. В ходе недавнего исследования, проведенного совместно группой Синтеза природных соединений и лабораторией Биомолекулярной ЯМР-спектроскопии ИБХ РАН в Москве, а также красноярской лабораторией фотобиологии ИБФ СО РАН, была расшифрована структура и проведен полный синтез нового люциферина, обладающего уникальными химическими свойствами. Этот люциферин является ключевым компонентом новой АТФ-зависимой биолюминесцентной системы.
Холодный свет жизни
Летом на Черном море и на Дальнем Востоке воздух ночами кишит живыми летающими фонариками, своды пещер Австралии и Новой Зеландии сияют ярче звездного неба, в тропиках бывают ночи, когда светится море (рис. 1). В сибирской тайге тоже можно столкнуться с подобным — земля иной раз мерцает под ногами, да только зрелище это не для любителей домашнего комфорта: нужно оказаться в лесу ночью в дождь, когда черви вылезают на поверхность, либо снять с поверхности почвы слой листового опада и травы, — вот тогда вы и увидите тысячи мелких ярких «звездочек».

Рисунок 1. Слева: Свечение мушек Arachnocampa luminosa в австралийской пещере. Структура люциферина этих насекомых до сих пор не установлена. Справа: Светящиеся черви Fridericia heliota на поверхности почвы в сибирской тайге. Открытый нами люциферин этих олигохет стал восьмым в ряду известных науке люциферинов, завершив
Размышляли ли вы о том, кто из живых существ умеет светиться, почему и как это происходит? Светятся, например, бактерии, кораллы, жуки, медузы, моллюски, черви, грибы, рыбы. Излучение света живыми организмами происходит благодаря химической реакции окисления небольшой органической молекулы — люциферина. В ней участвуют: окислитель (чаще всего кислород воздуха, иногда перекись водорода), восстановитель (люциферин) и фермент, называемый люциферазой. Только в трех известных случаях участником реакции также является АТФ (многоножка Luminodesmus sequoia, жуки и грибные мушки рода Arachnocampa). На данный момент известны структуры только семи природных люциферинов, последняя из которых была расшифрована более четверти века назад [1].
Явление биолюминесценции получило широкое практическое применение. Экологи используют его для мониторинга окружающей среды, в медицине и фармацевтике биолюминесценцию применяют для проведения клинических анализов и в тест-системах для поиска лекарств. В фундаментальных биохимических исследованиях биолюминесценция применяется для визуализации физиологических процессов, происходящих в клетках и целых организмах, а также для определения различных аналитов, в первую очередь — АТФ, ферментов, антител, антигенов. В генно-инженерных исследованиях самый надежный метод «увидеть» работу гена — «прицепить» к нему люциферазу и добавить к клеткам люциферин. Этот метод, например, был использован в недавней работе сотрудников ИБХ РАН про «молекулярное одомашнивание» ретровируса человека [2].
Открытие нового вида биолюминесцентных кольчатых червей семейства энхитреид — Fridericia heliota, обнаруженного в таежных районах юга Красноярского края, привело наш коллектив к структуре нового люциферина — компонента биолюминесцентной системы нового типа. Исследования проведены тремя научными группами: московским коллективом сотрудников группы синтеза природных соединений ИБХ РАН под руководством Ильи Ямпольского, Максимом Дубинным и Кириллом Надеждиным из лаборатории Биомолекулярной ЯМР-спектроскопии ИБХ РАН и группой Валентина Петушкова из красноярской лаборатории фотобиологии Института биофизики СО РАН.
Предыстория
Началось все с простого любопытства. Когда Валентин Петушков был еще студентом, работая ночью на биостанции Красноярского Университета, он обратил внимание на светящиеся точки в собственных следах на земле. Проверив горсть почвы, он заметил там небольших червей, испускающих голубоватое свечение. Поигрался с ними и забыл (как и многие студенты, думал — наверняка все тут уже давно известно и изучено, ведь ХХ век на дворе). А вспомнил об этом случае много позже, уже профессионально занимаясь бактериальной люминесценцией в лаборатории фотобиологии ИБФ СО РАН. Как-то на обеденном чаепитии во время разговора на околорабочие темы и прозвучал вопрос, повисший в воздухе: «А что за черви-то светились?»
Вот так, в конце
На тот момент было известно, что 12 видов из 6 родов олигохет имеют единый характер люминесценции: все эти черви выделяют целомическую жидкость, в клетках которой сосредоточена перекись-зависимая биолюминесцентная система. Наиболее полно она изучена у Diplocardia longa — крупных (~60 см, ~7 г) червей, обитающих в песчаных почвах юга Джорджии (США). Люциферин D. longa идентифицирован как алифатический альдегид N-изовалерил-З-амино-1-пропаналь и проявляет себя как субстрат в экстрактах целомической жидкости других светящихся олигохет. Но в случае с люциферазой Fridericia heliota результат кросс-реакции был отрицательным, что означало, что Fridericia использует другую биолюминесцентную систему.
В 2003 году вместе с итальянским специалистом по систематике червей Эмилией Рота было сделано полное подробное описание «светящегося червя из Сибирской тайги» [4].
В

Рисунок 2. Сбор таежной почвы в местах скопления червей F. heliota.
В 2011 году был выигран «мегагрант» на создание лаборатории биолюминесцентных биотехнологий в Сибирском Федеральном Университете, работу в которой возглавил выдающийся ученый, многие годы изучавший явление биолюминесценции, — нобелевский лауреат Осаму Шимомура [8]. Проект был посвящен фундаментальным исследованиям физико-химических механизмов биолюминесценции грибов, морских беспозвоночных и червей и позволил интенсифицировать работы по изучению биолюминесцентной системы червя F. Heliota (рис. 3).
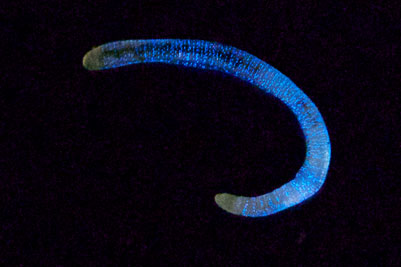
Рисунок 3. Люминесценция червя Fridericia heliota. Фотография любезно предоставлена Александром Семеновым (Беломорская Биологическая станция МГУ).
Путь к структуре люциферина
Путь к установлению структуры природного вещества не может обойтись без его выделения в чистом виде, и здесь группе Валентина Петушкова достался «счастливый» жребий собирать бесценную субстанцию годами по невидимым глазу крупицам, вручную выбирая из земли тысячи и тысячи светящихся фридериций. Ручной сбор в тайге давал около 30 г очищенной биомассы в год. Содержание люциферина во фридерициях составляет ~0.1 микрограмма на грамм необработанной биомассы, и в дополнение ко всему вышеперечисленному червяки совершенно не желают размножаться в лабораторных условиях. Из 90 г биомассы червей, собранных за несколько лет, группе Валентина Петушкова удалось выделить лишь 5 мкг люциферина. Помимо самого люциферина, экстракт F. heliota содержит несколько соединений неустановленного состава, обладающих схожими с люциферином спектральными и хроматографическими свойствами. Эти компоненты имеют похожий спектр и флуоресцируют так же, как и настоящий люциферин, но люциферином не являются. Валентин Петушков предположил, что эти соединения могут являться неактивными аналогами люциферина — его предшественниками или продуктами деградациии, а значит, они должны иметь схожую с люциферином структуру.
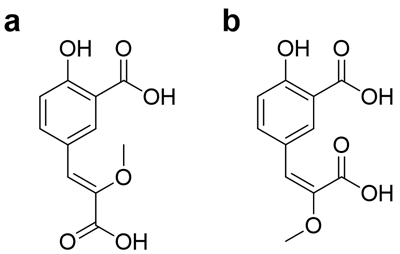
Рисунок 4. CompX — аналог люциферина червя F. Heliota. Структуры CompX (a) и его неприродного изомера (b).
Одним из основных компонентов экстракта червя было соединение, названное CompX, которого оказалось в 30 раз больше люциферина. Этого количества (~150 мкг) оказалось достаточно, чтобы на самом современном ЯМР-спектрометре получить всю информацию о его структуре. По данным ЯМР- и масс-спектров сотрудниками ИБХ Максимом Дубинным и Кириллом Надеждиным была предложена структура этого соединения. Молекула CompX, вероятнее всего, является необычным производным природной аминокислоты — тирозина, получаемым в результате трех модификаций: дезаминирования до кетокислоты, метилирования енола и карбоксилирования в ароматическое кольцо (рис. 4). Несмотря на кажущуюся простоту, молекулу с таким строением в природе еще не находили. Для установления конфигурации двойной связи в молекуле CompX необходимо было синтезировать и сравнить спектральные данные обоих изомеров; синтез был выполнен Александрой Царьковой и Михаилом Барановым. Z-изомер оказался идентичен природному образцу. Напротив, изомер CompX с противоположной конфигурацией двойной связи (Е-изомер) обладал значимо отличными спектральными свойствами, одним из которых было отсутствие флуоресценции.
В ходе дальнейших исследований биолюминесцентной системы червя методом ЯМР было обнаружено, что CompX является структурным фрагментом как самого люциферина, так и нескольких его аналогов, содержащихся в экстракте биомассы. Установление точных структур всех аналогов было необходимо для обеспечения более полного понимания путей биосинтеза и биодеградации люциферина, что привело к необходимости синтезировать эти соединения.
Что касается природного люциферина, чрезвычайно малое его количество (всего 5 мкг) позволило получить лишь масс-спектры высокого разрешения и только часть ЯМР-спектров, необходимых для установления структурной формулы. Самый современный ЯМР-спектрометр в России — Avance III 800 МГц фирмы Брукер с криогенно-охлажденным датчиком — позволил за неделю экспериментального времени накопить только протонный 1Н ЯМР-спектр, спектры COSY, HSQC и пустой (состоящий из шума криогенно-охлажденного датчика) спектр HMBC. Этот пустой спектр, если бы он содержал значимые сигналы, позволил бы соединить между собой различные фрагменты молекулы и получить целиком структурную формулу люциферина Fridericia heliota. Однако, вместо этого в руках исследователей остались только три фрагмента люциферина, расшифрованные по спектрам 1H, COSY и HSQC: остатки CompX (он был ранее расшифрован по полному набору ЯМР спектров), лизина и γ-аминомасляной кислоты (ГАМК). Данные масс-спектров позволили установить брутто-формулу люциферина: C23H29N3O11 и, сложив массы найденных в ЯМР фрагментов (за вычетом двух молекул воды, которые освобождаются при образовании двух пептидных связей), обнаружить «недостачу» с брутто-формулой люциферина в количестве C2O3.
«Недостача» как раз соответствует остатку щавелевой кислоты (оксалата), которая невидима в тех ЯМР-экспериментах, которые удалось накопить с 5 мкг вещества. Таким образом стало очевидно, что молекула люциферина образована четырьмя фрагментами: CompX, лизин, ГАМК и оксалат. Итак, самая правдоподобная гипотеза о химической природе люциферина F. heliota — это необычный олигопептид, состоящий из четырех остатков. Существует всего 10 способов собрать олигопептид из этих четырех фрагментов, соединяя пептидной связью карбоксильные и амидные группы. Группа синтеза природных соединений под руководством Ильи Ямпольского могла бы синтезировать все 10 вариантов, если бы потратила на это год работы, а группа ЯМР-спектроскопии могла бы накопить HMBC спектр люциферина, если бы потратила несколько месяцев ценнейшего приборного времени исключительно на регистрацию одного ЯМР-спектра (ведь снимали же на БАКе бозон Хиггса два года?). Сэкономить приборное время и время химиков-синтетиков помог Максим Дубинный: ему удалось отличить «свободные» карбоксильные группы в пептидной связи от «занятых», сняв всего 10 1H ЯМР-спектров при разных значениях рН, что заняло один день приборного времени. Дело в том, что «свободные» карбоксильные группы титруются при изменении рН: переходят из формы COOH в кислом растворе в фому COO− в нейтральной и щелочной среде, и эти изменения отражаются на соседних группах, за которыми можно наблюдать методом ЯМР.
Данные по титрованию показали, что обе карбоксильные группы CompX участвуют в образовании пептидной связи, а карбоксилы лизина и ГАМК свободны. Теперь спектральным данным не противоречили всего четыре изомерных структуры, которые различались порядком пептидных связей, соединяющих фрагменты (рис. 5). На этом возможности спектральных методов закончились, и выбирать, какой из четырех возможных изомеров отражает строение люциферина, пришлось группе синтеза природных соединений полным синтезом теперь уже четырех оставшихся изомеров. Такой синтез был предпринят, и только синтетическое соединение 1 оказалось идентичным по всем спектральным характеристикам природному люциферину и, что самое важное, оно вступило в реакцию биолюминесценции с люциферазой червя с испусканием света (рис. 6). «Момент истины» настал 14 октября 2013 года: в этот день были получены спектры ЯМР, идентичные спектрам природного люциферина, а спустя два часа зарегистрирован сигнал синтетического люциферина на люминометре в присутствии АТФ и белкового экстракта червя.
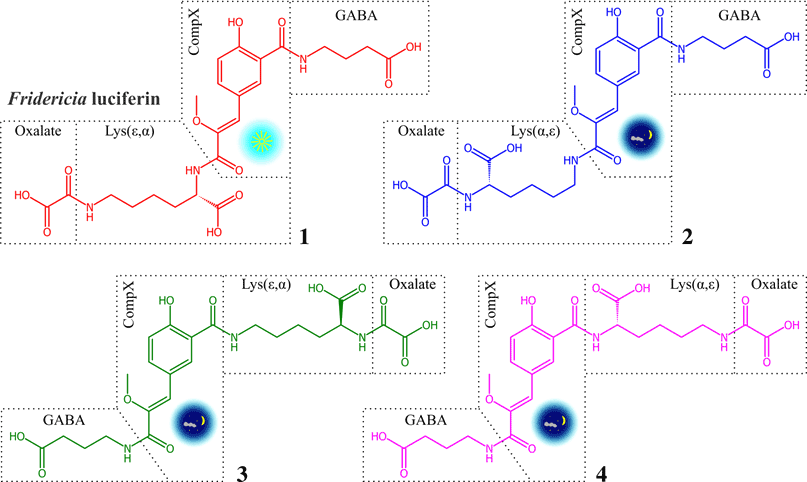
Рисунок 5. Структуры четырех пептидных изомеров люциферина. Только соединение 1 испускало свет при смешении с люциферазой F. heliota.

Рисунок 6. Свечение синтетического люциферина Fridericia в присутствии АТФ и белкового экстракта червя.
Что же дальше?
В результате успешной коллаборации трех научных групп из сибирского почвенного червя Fridericia heliota нами выделен новый люциферин — ключевой компонент новой АТФ-зависимой биолюминесцентной системы. Мы установили его структуру и провели встречный синтез. Возможно, механизм биолюминесценции подобен химической реакции, протекающей в хемилюминесцентных «светящихся палочках». Вероятно, в ходе реакции люминесценции происходит окисление одной из трех свободных карбоксильных групп люциферина, в то время как флуоресцентный остаток CompX отвечает за испускание квантов света. Результаты исследований опубликованы в журналах Tetrahedron Letters и Angewandte Chemie
Открытый нами люциферин Friericia heliota имеет шансы занять свою нишу в области прикладной биолюминесценции. Во-первых, он прост в химическом синтезе: группа профессиональных химиков сделает себе такой же примерно за месяц. Во-вторых, он исключительно стабилен: при комнатной температуре в водном растворе сохраняет свою активность в течение месяцев, и даже лет. Люциферин светлячков — наиболее популярный сегодня люциферин в лабораторных и промышленных анализах — теряет свою активность в водном растворе уже в течение нескольких часов. В-третьих, в отличие от люциферина бактерий, он не токсичен.
На данный момент основное направление наших совместных исследований — структура различных аналогов люциферина и их роль в его биосинтезе (метаболизме), структура продукта окисления люциферина (оксилюциферин), установление аминокислотной последовательности и рекомбинантная продукция люциферазы — пока еще неизвестного нам фермента, который катализирует реакцию биолюминесценции. В дальнейшем мы планируем установить пути биосинтеза люциферина, выделить все ответственные за это ферменты и кодирующие их гены и перенести биолюминесценцию из олигохет в бактерию, а потом — очень может быть — и в подходящий эукариотический организм.
Текст написан коллективом авторов работы. Основные результаты опубликованы в журнале Angewandte Chemie [9].
Литература
-
Nakamura H., Kishi Y., Shimomura O., Morse D., Hastings J.W. (1989). Structure of dinoflagellate luciferin and its enzymic and nonenzymic air-oxidation products. J. Am. Chem. Soc. 111,
7607–7611; - Элементы: «Вирус, встроившийся в геном наших предков, влияет на работу мозга»;
-
Zalesskaya N.T., Petushkov V.N., Rodionova N.S. (1990). Dokl. Akad. Nauk SSSR 310(2),
496–498 ; -
Rota E., Zalesskaja N.T., Rodionova N.S., Petushkov V.N. (2003). Redescription of Fridericia heliota (Annelida, Clitellata: Enchytraeidae), a luminous worm from the Siberian taiga, with a review of bioluminescence in the Oligochaeta. J. Zoology 260,
291–299. - Petushkov V.N., Rodionova N.S., Bondar V.S. (2003). Study of the Luminescence System of the Soil Enchytraeid Fridericia heliota (Annelida: Clitellata: Oligochaeta: Enchytraeidae). Dokl. Biochem. Biophys. 391, 204;
- Rodionova N.S., Bondar V.S., Petushkov V.N. (2003). ATP Is a Cosubstrate of the Luciferase of the Earthworm Fridericia heliota (Annelida: Clitellata: Oligochaeta: Enchytraeidae). Dokl. Biochem. Biophys. 392, 253;
-
Petushkov V.N., Rodionova N.S. (2007). Purification and partial spectral characterization of a novel luciferin from the luminous enchytraeid Fridericia heliota. J. Photochem. Photobiol. B 87,
130–136; - биомолекула: «Флуоресцирующая Нобелевская премия по химии»;
- Petushkov V.N., Dubinnyi M.A., Tsarkova A.S., Rodionova N.S., Baranov M.S., Kublitski V.S., Shimomura O., Yampolsky I.V. (2014). A novel type of luciferin from Siberian luminous earthworm Fridericia heliota: structure elucidation by spectral studies and total synthesis. Angew. Chem., Int. Ed. doi: 10.1002/anie.201400529;
-
Petushkov V.N., Dubinnyi M.A., Rodionova N.S., Nadezhdin K.D., Marques S.M., Esteves da Silva J.C.G., Shimomura O., Yampolsky I.V. (2014). AsLn2, a luciferin-related modified tripeptide from the bioluminescent earthworm Fridericia heliota, Tetrahedron Lett. 55,
463–465; -
Petushkov V.N., Tsarkova A.S., Dubinnyi M.A., Rodionova N.S., Marques S.M., Esteves da Silva J.C.G., Shimomura O., Yampolsky I. V. (2014). CompX, a luciferin-related tyrosine derivative from the bioluminescent earthworm Fridericia heliota. Structure elucidation and total synthesis. Tetrahedron Lett. 55,
460–462.
16 апреля 2014 года
Источник: http://onlinelibrary.wiley.com/doi/10.1002/anie.201400529/abstract